
Research focus
This requires a detailed knowledge of which nerve cells take part, how they talk to each other through synaptic interaction and an understanding of the intrinsic function of these networks.
Essentially our research extends from ion channels and synapses to network mechanisms and behaviour utilising a multitude of techniques from patch clamp, tract tracing and cellular imaging to modelling and studies of behaviour. We utilise preferentially the lamprey as model organism.
We have been able to successfully model, based on detailed cellular knowledge, the networks responsible for the command and pattern generating systems for locomotion including steering and posture.
Our work continues with several foci including the role of the pallium/cortex, basal ganglia for selection of different patterns of motor behaviour, optic tectum for steering and eye motor coordination, the physiological role of different modulator systems acting through the spinal networks, and different ion channel subtypes contributing to neuronal function.
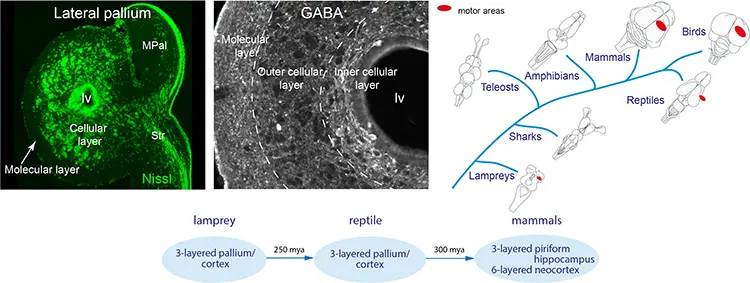
Our recent findings have shown that the lamprey forebrain has all components of the mammalian forebrain – a finding that has radically changed the view on the evolutionary origin of the vertebrate forebrain. The basic organisation had evolved 560 rather than 300 million years ago as previously believed.
The projects are supported by The Swedish Research Council, EU (Human Brain Project) and Karolinska Institutet.
Evolutionary conservation of the lamprey forebrain
Cortex/pallium is three-layered with the same efferent projection pattern to the brainstem-spinal cord as in mammals.
The lamprey pallium/cortex is three-layered with an inner and outer granular layer and a molecular layer. The GABAergic cells are most abundant in the inner granular layer. Phylogenetically, a layered pallium/cortex has been demonstrated in mammals, birds, reptiles and the lamprey.
Publications
- Suryanarayana SM, Robertson B, Grillner S. (2022) The neural bases of vertebrate motor behaviour through the lens of evolution. Philos Trans R Soc Lond B Biol Sci 377(1844):20200521
- Suryanarayana SM, Pérez-Fernández J, Robertson B, Grillner S. (2021) The Lamprey Forebrain - Evolutionary Implications. Brain Behav Evol. 30:1-16
- Suryanarayana SM, Robertson B, Wallén P, Grillner S (2017) Curr Biol 27:1-14.
- Ocana F, Suryanarayana SM, Saitoh K, Kardamakis AA, Capantini L, Robertson B, Grillner S. (2015) Curr Biol 25:1-11
Sensory representation in the lamprey pallium/cortex.
Visual, somatosensory and motor areas in the dorsal pallium and olfactory representation in the ventral pallium are a general and common schema of vertebrate pallial organisation.
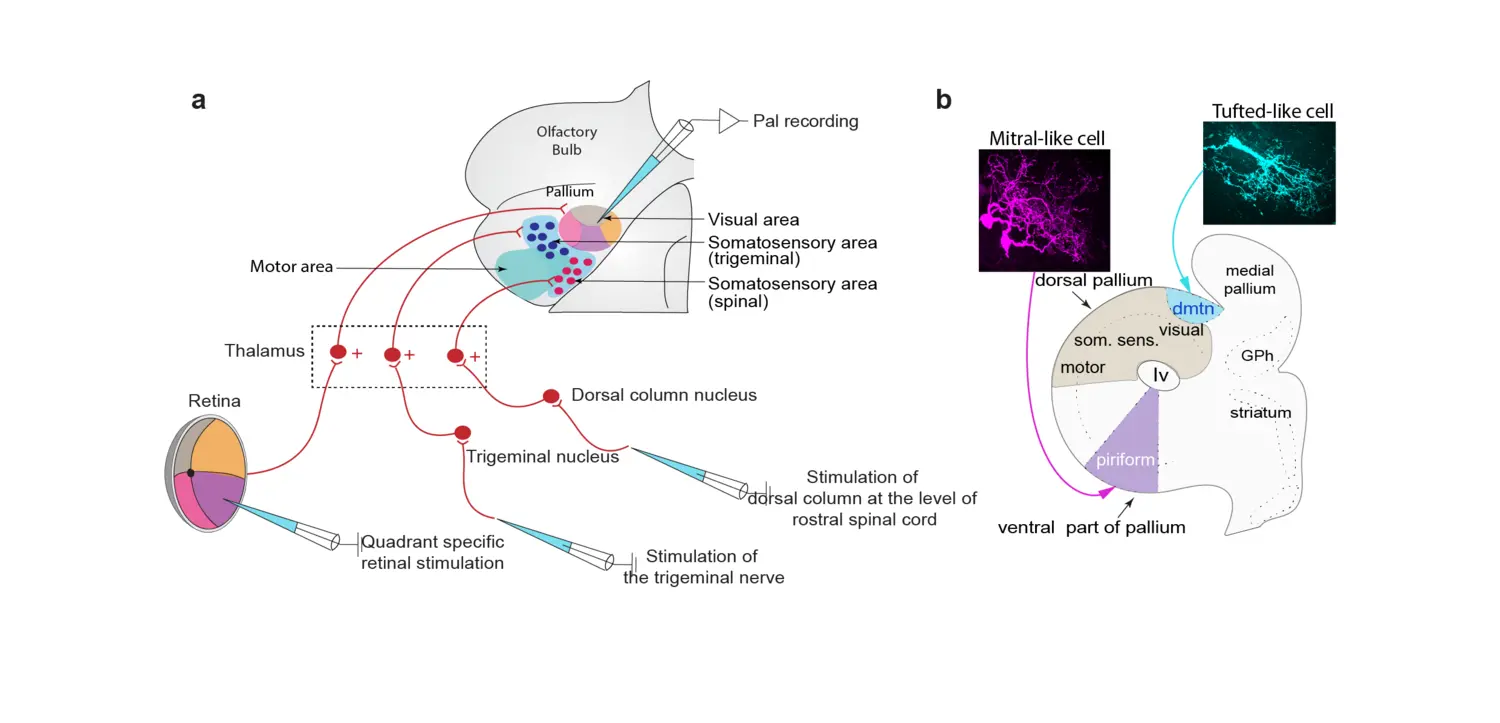
a. Summarising schematic of the lamprey dorsal pallium, showing retinotopic visual areas, somatosensory areas and motor areas, as well as the retinal, trigeminal and dorsal column nucleus afferents relayed via distinct subpopulations of thalamic neurons.
b. The olfactory system in lamprey resembles that of mammals in many respects with a dual efferent system conveyed via tufted- and mitral-like cells. The latter target the olfactory cortex located in the ventral pallium, whereas the former target a separate limited region, the dmtn (dorsomedial telencephalic nucleus). The mitral-like (magenta) and tufted-like (blue) cells display different morphology.
Publications
- Suryanarayana SM, Pérez-Fernández J, Robertson B, Grillner S. (2021) Cell Rep. 34:108596
- Suryanarayana SM, Pérez-Fernández J, Robertson B, Grillner S. (2020) Nat Ecol Evol. 4:639-651
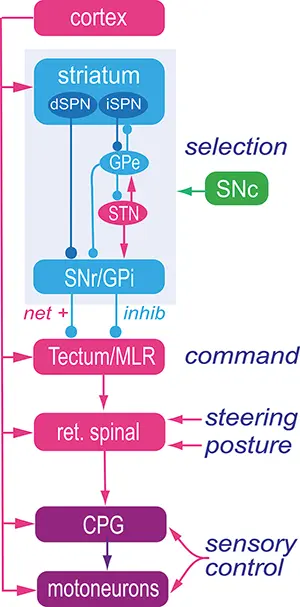
The lamprey basal ganglia and the dopamine system is conserved
The organisation of the basal ganglia including the dopamine system is virtually identical throughout vertebrate phylogeny – from lamprey to primates. This applies to the overall neural organisation, transmitters, peptides, synaptic connectivity and expression of ion channels.
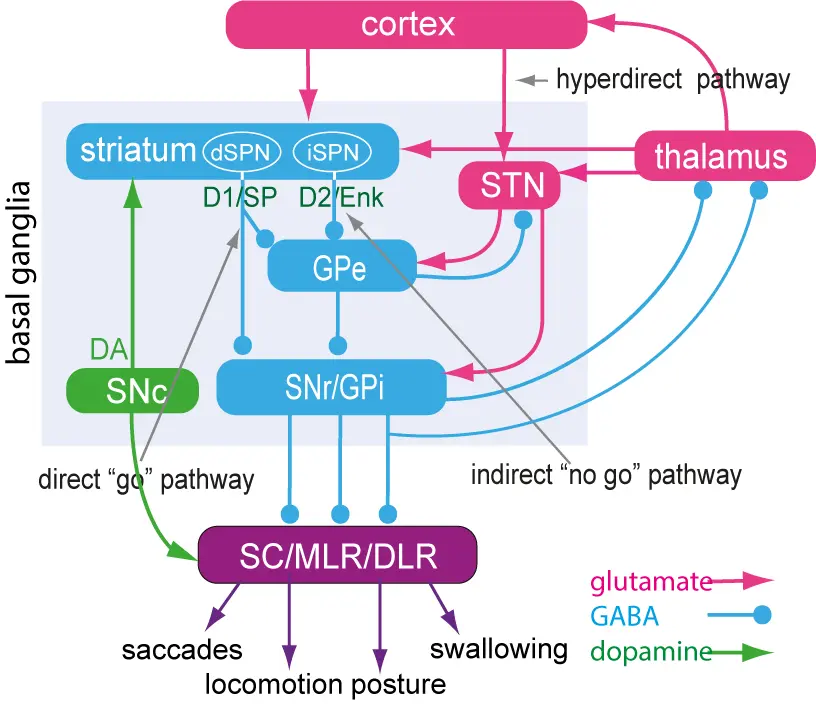
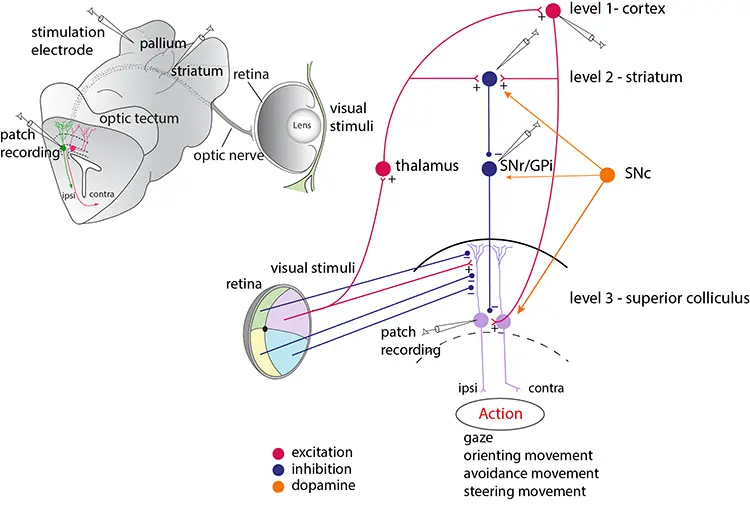
The striatum consists of GABAergic neurons as do globus pallidus externa (GPe), globus pallidus interna (GPi) and substantia nigra pars reticulata (SNr). SNr and GPi represent the output level of the basal ganglia, which project via different subpopulations of neurons to the optic tectum (superior colliculus), the mesencephalic (MLR) and diencephalic (DLR) locomotor command regions and additional brainstem motor centres, and also back to thalamus with efference copies of information sent to the brainstem. The indirect loop is represented by the GPe, the subthalamic nucleus (STN) and the output level (SNr/GPi) — the net effect being an enhancement of activity in these nuclei. The striatal neurons of the direct pathway to SNr/GPi express the dopamine D1 receptor (D1) and substance P (SP), while the indirect pathway neurons in striatum express the dopamine D2 receptor (D2) and enkephalin (Enk). Also indicated is the dopamine input from the substantia nigra pars compacta (SNc, green) to striatum and brainstem centres.
Publications
- Grillner S. Proc Natl Acad Sci U S A. 2025 Apr;122(13):e2423068122. doi: 10.1073/pnas.2423068122
- Frost-Nylén J, Thompson WS, Robertson B, Grillner S. Curr Neuropharmacol. 2024;22(9):1419-1430
- Grillner S, Robertson B, Kotaleski JH. (2020) Compr Physiol. 10:1241-1275
- Grillner S and Robertson B. (2016) Curr Biol, 26: R1088-1100
- Pérez-Fernández J, Kardamakis AA, Suzuki DG, Robertson B, Grillner S. (2017) Neuron 96:910-924.e5.
- Stephenson-Jones M, Samuelsson E, Ericsson J, Robertson B and Grillner S (2011) Curr. Biol. 21:1081-91
The interaction between the forebrain and the optic tectum
The optic tectum contains motor circuits for the control of eye and orienting/evasive movements and receives visual input arranged in a retinotopic map. The optic tectum is controlled from both the basal ganglia and cortex/pallium. We have used an isolated eye-brain preparation to allow for a detailed analysis.
Publications
- Wibble T, Pansell T, Grillner S, Pérez-Fernández J. Nat Commun. 2022 Aug 10;13(1):4699
- Isa T, Marquez-Legorreta E, Grillner S, Scott EK (2021) The tectum/superior colliculus as the vertebrate solution for spatial sensory integration and action. Curr Biol 31(11):R741-R762
- Pérez-Fernández J, Kardamakis AA, Suzuki DG, Robertson B, Grillner S. (2017) Neuron 96:1-15.
- Kardamakis A, Pérez-Fernández J, Grillner S. (2016) ELife pii: e16472. doi: 10.7554/eLife.16472.
- Kardamakis AA, Saitoh K. and Grillner S. (2015) Proc Natl Acad Sci USA E1956-65.
Modelling, the neural control of action
The mechanisms underlying the operation of the forebrain and midbrain in action selection are being investigated through detailed modelling based on experimental data, on the systems level as well as on the cellular and subcellular levels.
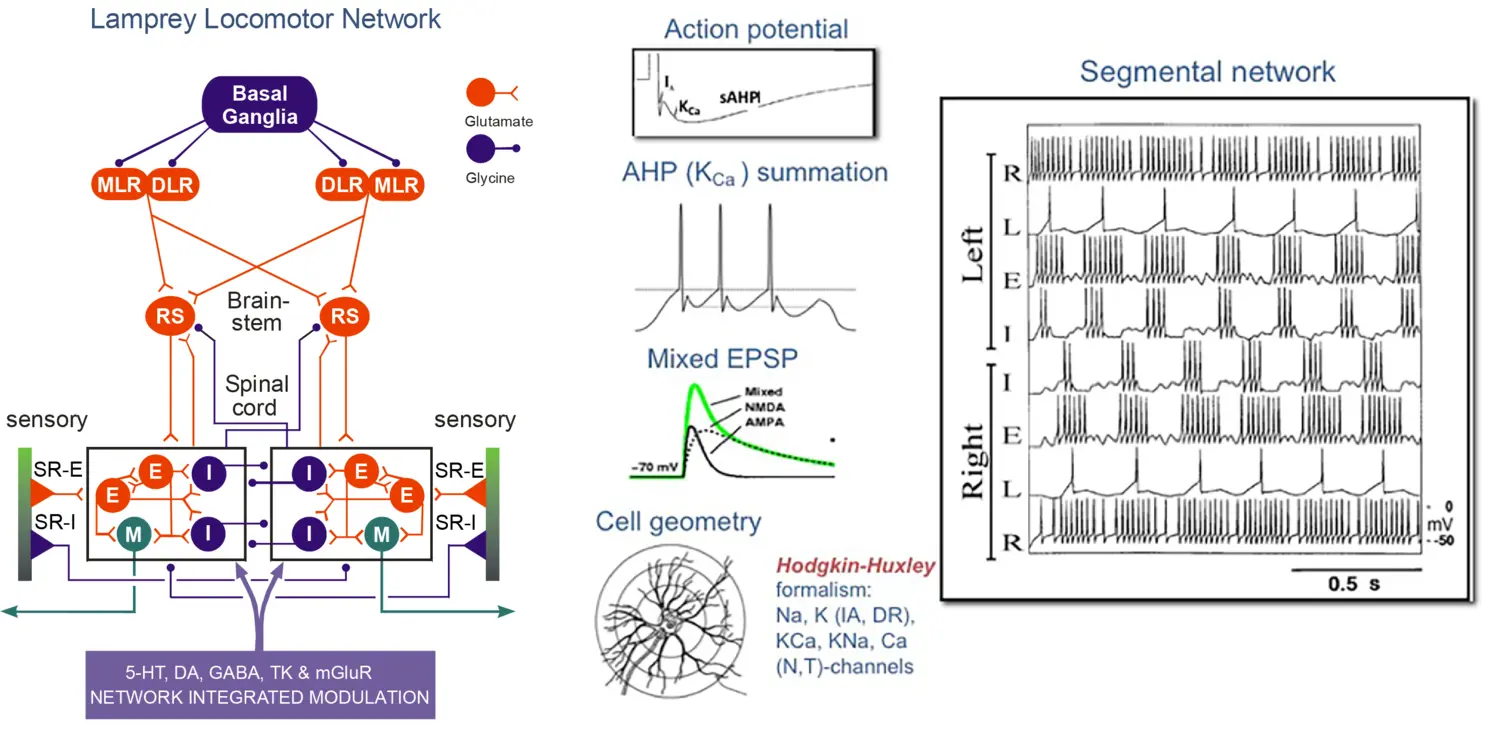
Locomotor network of the lamprey. Left: Schematic representation of the forebrain, brainstem and spinal components of the neural circuitry generates rhythmic locomotor activity. Middle: Neuron model simulation using Hodgkin-Huxley formalism of the action potential with fast and slow afterhyperpolarisation (AHP) and synaptic properties. Right: Network model simulation of rhythmic, alternating activity in the spinal segmental network.
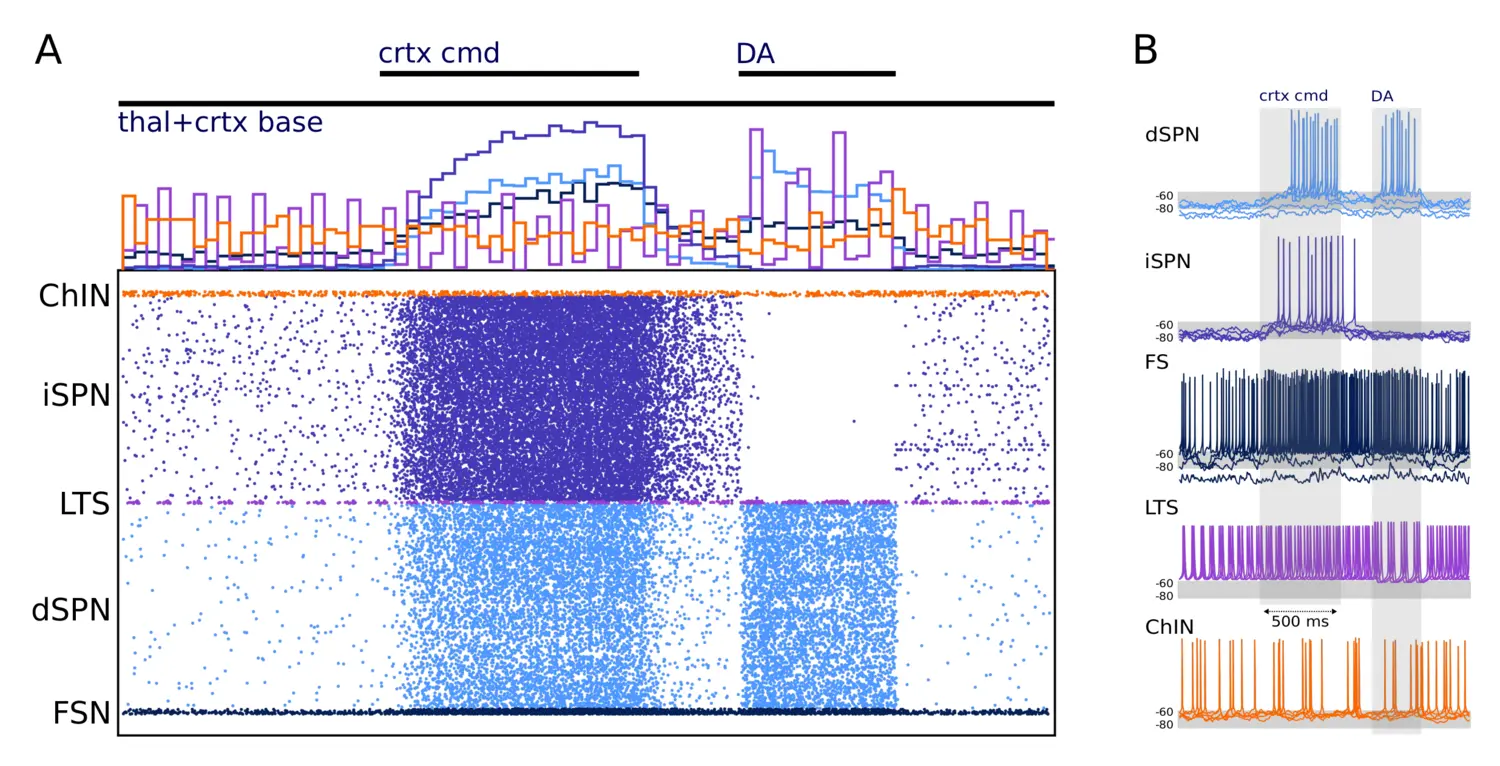
Network simulation of 10,000 neurons. (A) The activity of the network is shown in the form of a raster plot (Bottom) and spike histogram (Top). (B) Example traces of each cell type in the network are shown. The network is driven with cortical and thalamic input and modulated by dopamine, as indicated at the Top of the figure and the shaded areas (in A and B, respectively). The three inputs represent 1) baseline activation of cortical and thalamic input (thal+crtx baseline), 2) a cortical command signal (crtx cmd), during which the cortical activation is increased (given to all cells except the ChINs), and 3) a dopaminergic modulation signal that acts on conductances in accordance.
Publications
- Frost Nylén J, Hjorth JJJ, Kozlov A, Carannante I, Hellgren Kotaleski J, Grillner S. Proc Natl Acad Sci U S A. 2023 Nov 7;120(45):e2313058120
- Frost Nylén J, Carannante I, Grillner S, Hellgren Kotaleski J. (2021) Eur J Neurosci. 53:2135-2148
- Hjorth JJJ, Kozlov A, Carannante I, Frost Nylén J, Lindroos R, Johansson Y, Tokarska A, Dorst MC, Suryanarayana SM, Silberberg G, Hellgren Kotaleski J, Grillner S. (2020) Proc Natl Acad Sci U S A. 117:9554-9565
- Suryanarayana SM, Hellgren Kotaleski J, Grillner S, Gurney KN. (2019) Neural Networks 109:113-36
- Lindroos R, Dorst MC, Du K, Filipović M, Keller D, Ketzef M, Kozlov AK, Kumar A, Lindahl M, Nair AG, Pérez-Fernández J, Grillner S, Silberberg G, Hellgren Kotaleski J. (2018) Front Neural Circuits 12:3. doi: 10.3389/fncir.2018.00003. eCollection.
- Kozlov A, Kardamakis A, Hellgren Kotaleski J, Grillner S (2014) Proc Natl Acad Sci USA 111:3591-6
- Kozlov A, Huss M, Lansner A, Hellgren Kotaleski J, Grillner S (2009) Proc Natl Acad Sci USA 106:20027-32
- Grillner S, Cangiano L, Hu G, Thompson R, Hill R, Wallén P (2000) Brain Res 886:224-36.
The enigmatic cerebrospinal fluid-contacting neurons sense deviations from normal pH at the central canal and in hypothalamus
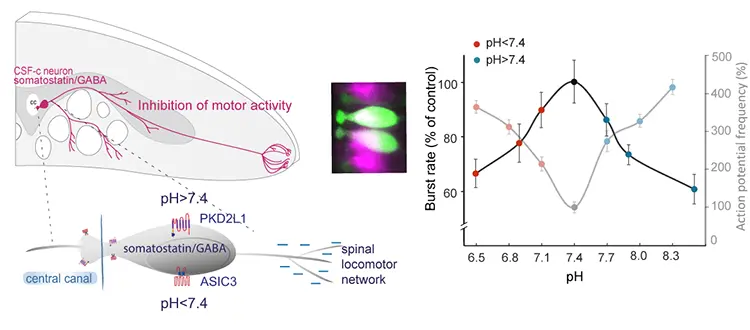
Both spinal and hypothalamic cerebrospinal fluid-contacting (CSF-c) neurons serve as pH sensors, thereby providing a novel homeostatic module for the regulation of pH in the CNS. In the spinal cord, acidic pH is mediated by the ASIC3 and alkaline pH via PKD2L1 channels, whereas in hypothalamus the acidic response is mediated via ASIC3 and alkaline pH via connexin hemichannels.
Publications
- Jalalvand E, Alvelid J, Coceano G, Edwards S, Robertson B, Grillner S, Testa I. (2022) ExSTED microscopy reveals contrasting functions of dopamine and somatostatin CSF-c neurons along the lamprey central canal. Elife Feb 1;;11:e73114.
- Jalalvand E, Robertson B, Tostivint H, Wallén P, Grillner S. (2018) J Neurosci. 38:7713-24.
- Jalalvand E, Robertson B, Wallén P, Grillner S. (2016) Nature Commun 7:10002. doi: 10.1038/ncomms10002.
